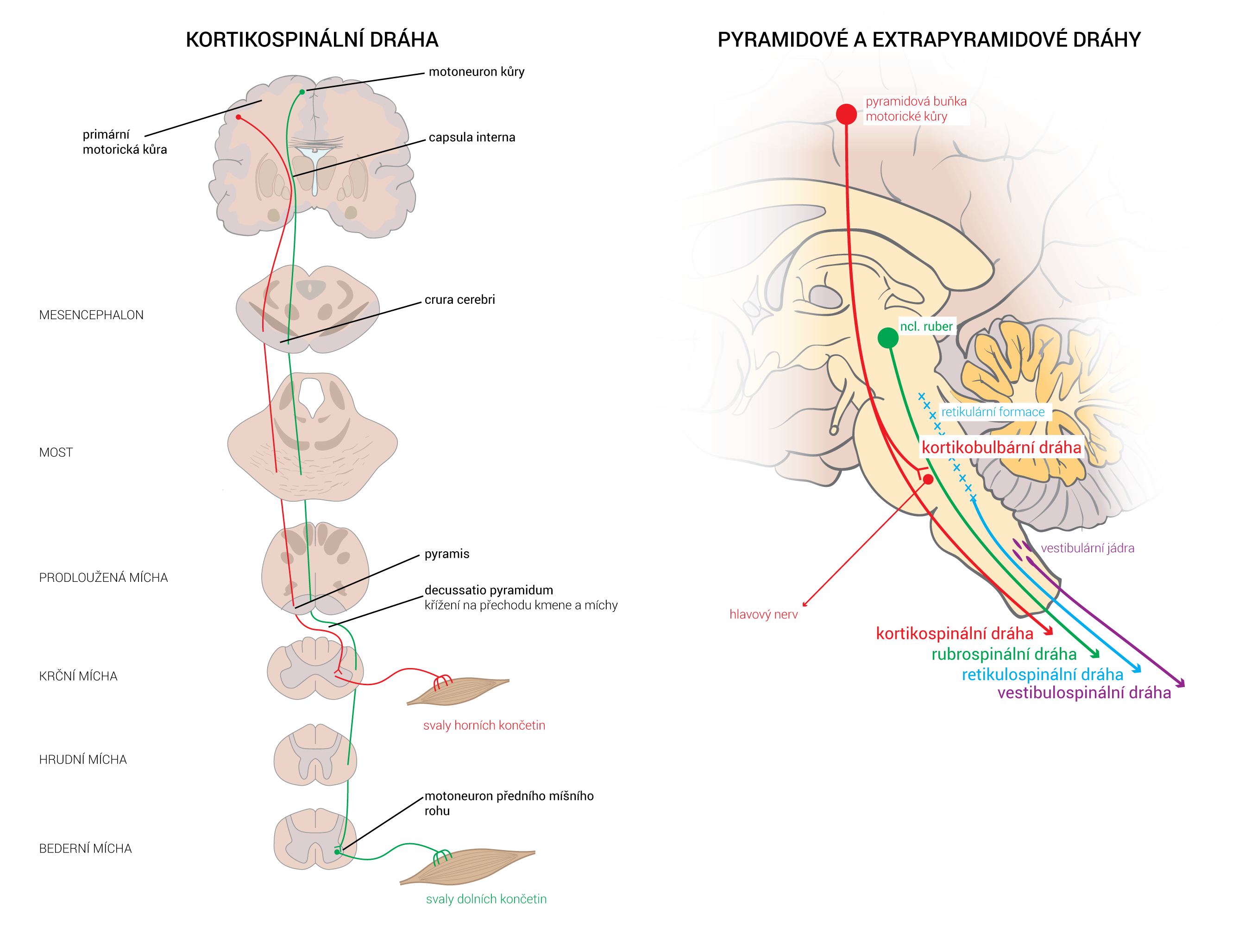
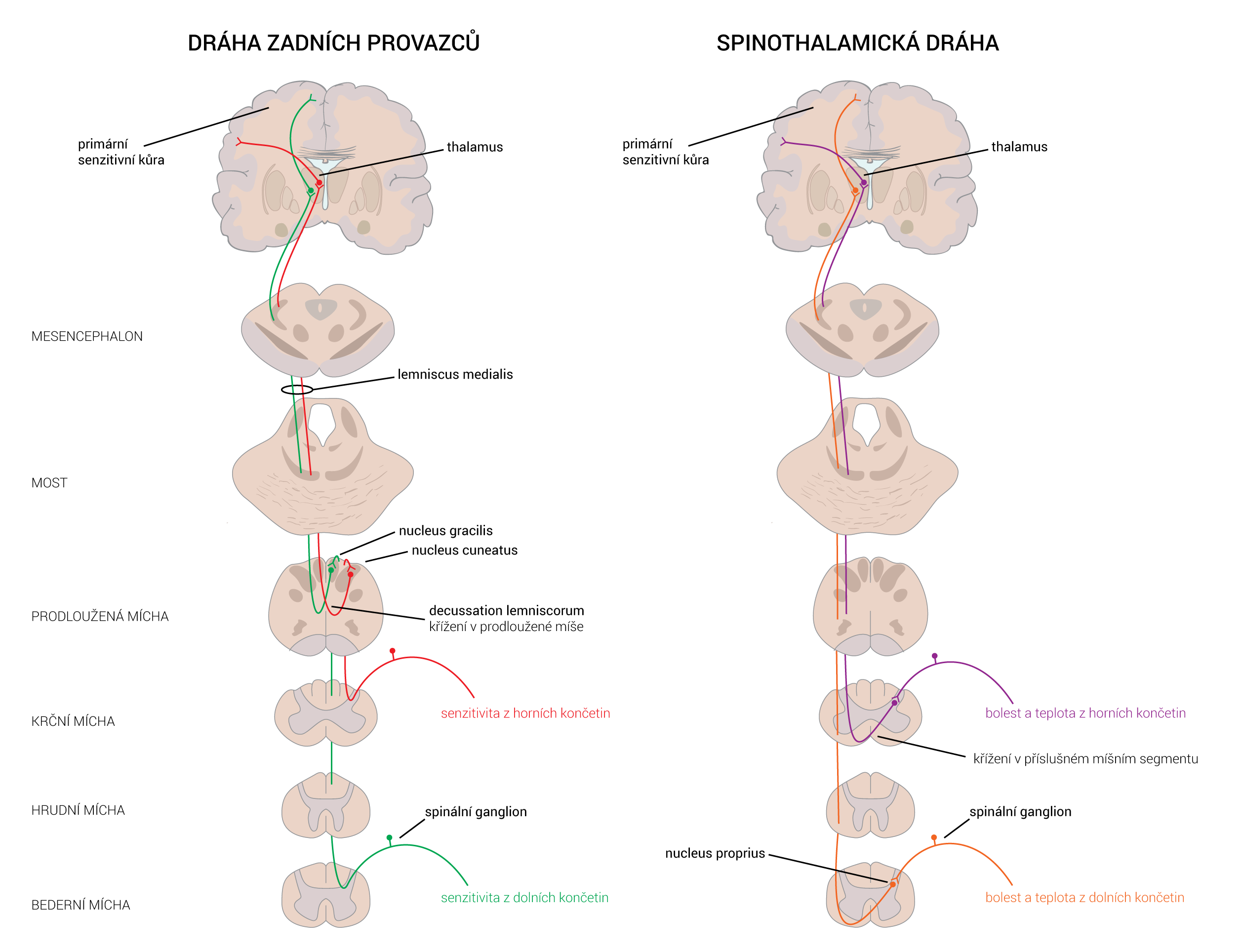
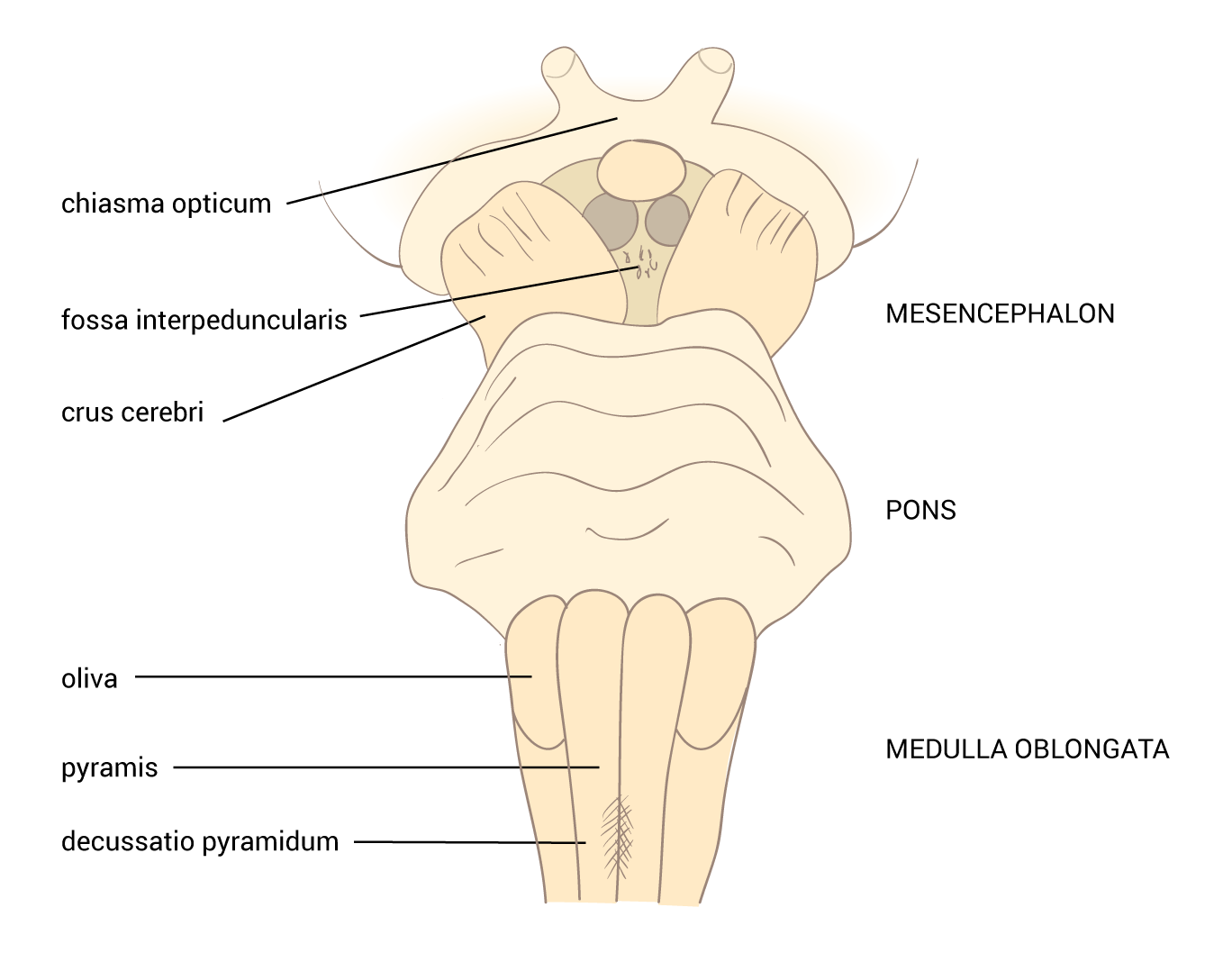
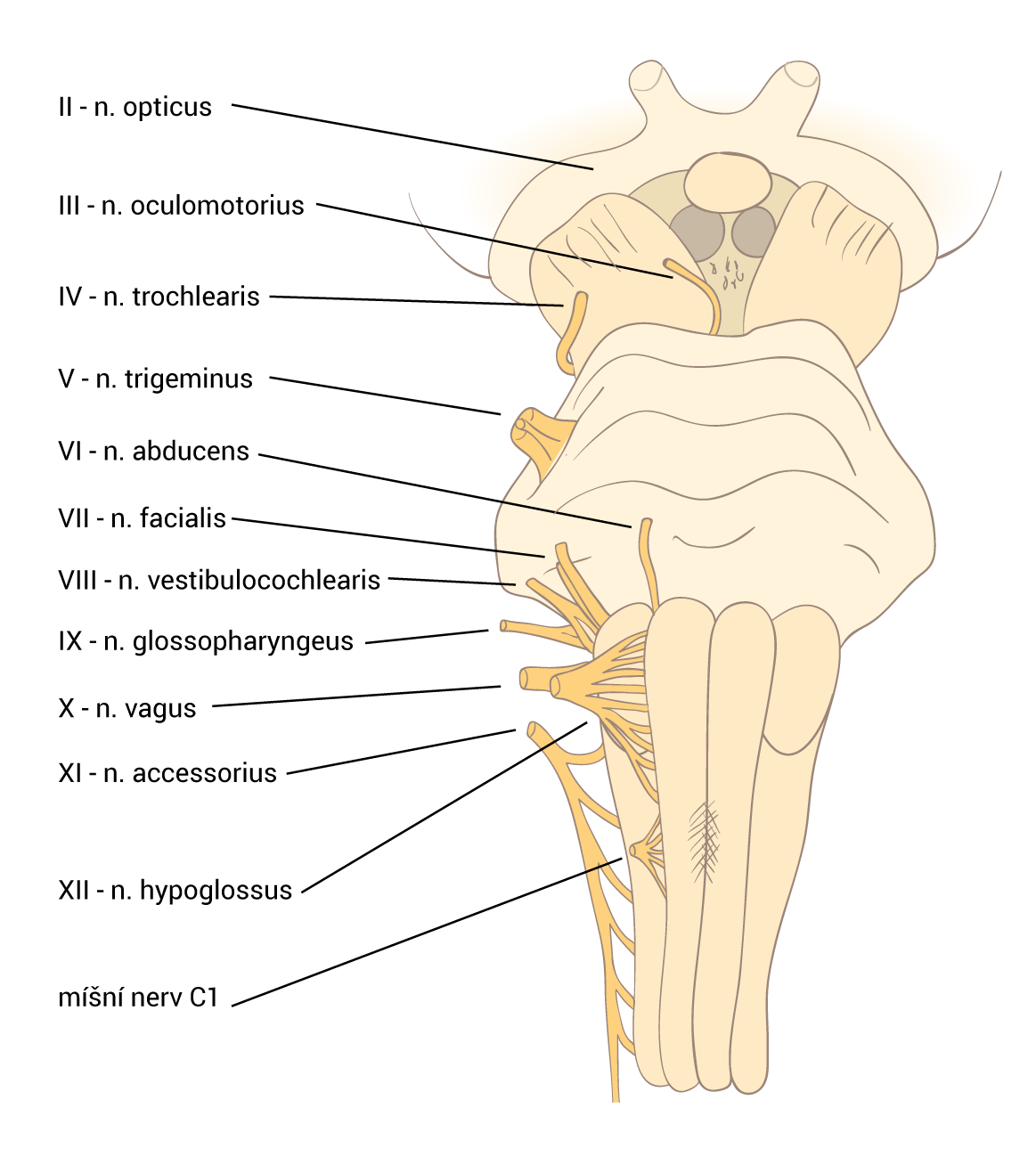
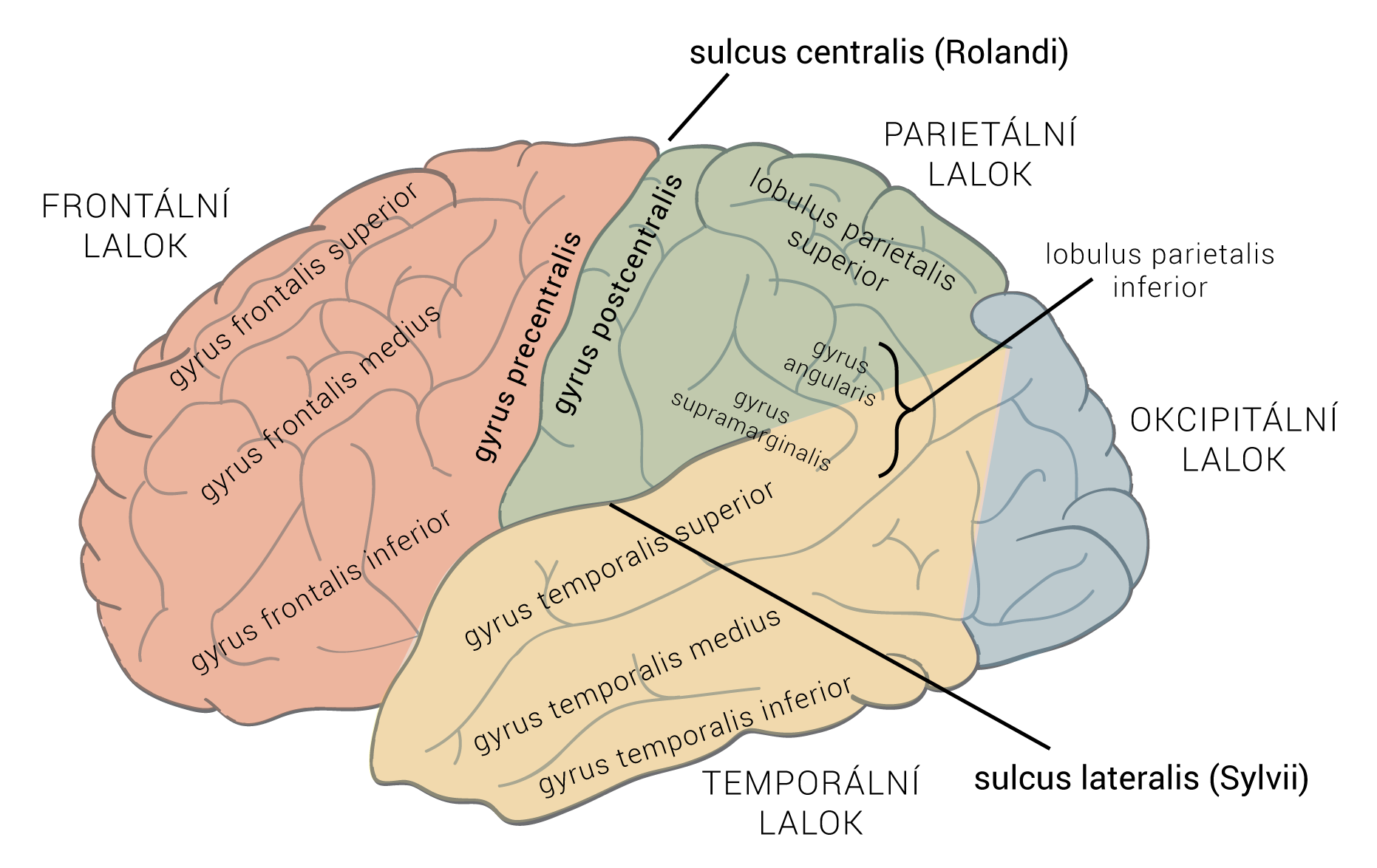
Neurotransmiter je látka uvolněná neuronem ke specifickým cílovým buňkám, aby v nich vyvolala specifický účinek. Cílové buňky mohou představovat jiné neurony či orgány (žlázy a svaly). Oproti endokrinní signalizaci, jež využívá k přenosu signálních molekul krevní oběh, neurotransmise využívá prosté difúze – signální molekuly jsou dopraveny axonálním transportem až do těsné blízkosti cílové buňky. Zde jsou uloženy ve vezikulách, ze kterých se uvoňují do synaptické štěrbiny.
Neurotransmitery se váží na receptory na postsynaptické membráně. Rychlé a krátkodobé změny jsou podmíněny ionotropními receptory, pomalé a dlouhodobé změny vyvolávají receptory spřažené s G-proteiny a receptory spřažené s enzymovou aktivitou.
Definice neurotransmiteru
Čtyři základní pravidla definují neurotransmitery proti ostatním látkám produkovaným v CNS:
1) látka se tvoří v presynaptickém neuronu
2) látka se skladuje v presynaptickém terminálu, ze kterého se exocytózou uvolňuje v dostatečném množství pro vyvolání odpovědi cílové buňky
3) exogenní podání látky vyvolá stejnou odpověď jako její endogenní uvolnění
4) účinek dané látky v synaptické štěrbině může být specificky inhibován
Ze strukturního pohledu jsou neurotransmitery velmi různorodou skupinou látek, následující dělení představuje jen jednu možnost jejich klasifikace.
- Malé molekuly
a) aminokyseliny: glutamát, aspartát či glycin
b) deriváty aminokyselin: GABA, katecholaminy – noradrenalin a dopamin, serotonin
c) acetylcholin
d) ostatní: puriny (ATP, ADP, adenosin), plyny (NO)
- Velké molekuly
a) peptidy: beta-endorfin, leu-enkefalin, substance P
b) endogenní kanabinoidy
Glutamatergní systém
Glutamát (Glu) je hlavní excitační neurotransmiter v CNS. Vyskytuje se celém CNS, nelze tedy uvést určitá specifická centra jeho tvorby či jeho projekce. Uplatňuje se v procesech vzniku nových synapsí, posílení funkčních synapsí i v zániku těch nevyužívaných – hovoříme o tzv. synaptické plasticitě, klíčovém mechanismu paměti a učení.
Tvorba a inaktivace glutamátu
Téměř pro všechny neurotransmitery platí fakt, že hematoencefalická bariera je špatně propustná pro funkční (hotové) neurotransmitery, a naopak skrze ní dobře prostupují jejich prekurzory. To stejné platí i pro glutamát; ten téměř nikdy neproniká z krve do CNS. Glutamatergní neurony produkují glutamát převážně pomocí enzymu glutaminázy z glutaminu. Glutamin vzniká v astrocytech prostřednictvím glutaminsyntetázy z glutamátu poté, co je jimi vychytán ze synaptické štěrbiny. Veškerá tvorba i inaktivace glutamátu tak probíhá v CNS touto interakcí neuron-astrocyt.
Receptory pro glutamát
Receptory pro glutamát se dělí podle svých agonistů:
1) AMPA-receptory
Ionotropní receptory, po otevření dochází k influxu Na+ a Ca2+ a efluxu K+.
2) NMDA-receptory
Ionotropní receptory umožňující průchod Na+, Ca2+ a K+. Ke svému otevření vyžadují jak vazbu ligandu, tak i určitou depolarizaci membrány.
Další typy zahrnují ionotropní kainátové receptory a skupinu metabotropních receptorů.
GABAergní systém
Kyselina gamma-aminomáselná (GABA) je hlavním inhibiční neurotransmiterem v CNS. Opět se vyskytuje povšechně.
Tvorba a inaktivace GABA
GABA se tvoří dekarboxylací glutamátu za katalýzy enzymem glutamátdekarboxylázou. I zde je hlavním prekurzorem glutamin transportovaný do GABAergních neuronů z astrocytů – dochází k přeměna glutamin – glutamát a následné dekarboxylaci. Ze štěrbiny je GABA vychytávána do astrocytu, kde se přes Krebsův cyklus přeměnuje na glutamin.
Receptory pro GABA
1) GABAA
Ionotropní receptor umožňující průchod Cl- do buňky.
2) GABAB
Metabotropní receptor spřažený s Gi-proteinem – inhibuje adenylátcyklázu, to vede k redukci intracelulárního cAMP, zvýšení permeability membrány pro K+ a oslabení přenosu skrze Ca2+ kanály. Výsledkem je inhibice neuronu.
3) GABAC
Ionotropní receptor spřažený s Cl- kanálem, vykazuje pomalejší otevírání a současně delší účinek oproti GABAA
Receptor GABAA je součástí tzv. supramolekulárního receptorového komplexu: kromě GABAA je tvořeni i receptory pro benzodiazepiny, barbituráty, kortikosteroidy a alkohol – všechny ligandy působí inhibici postsynaptického neuronu (přímé otevření Cl- kanálu, či zesílení působení GABA).
Cholinergní systém
Neurotransmiterem je acetylcholin (ACh). Mimo CNS působí na nervosvalových ploténkách, v pregangliových neuronech v autonomním nervovém systému i v postgangliových neuronech parasympatiku. V lidském mozku se uplatňuje v procesech paměti a učení a cyklu spánek-bdění.
Cholinergní neurony se vyskytují převážně v nc. basalis Meynerti a v jádrech septa – vysílají axony do kůry a hipokampální formace – aktivují kůru a uplatňují se v procesech učení a paměti, jejich poškození hraje roli při rozvoji Alzheimerovy choroby. Cholinergní neurony v tegmentu kmene vysílají axony do mozečku a hypotalamu – modulují cyklus spánek-bdění
– cholinergní neurony ve striatu se zapojují v okruhu bazálních ganglií – kontrola postoje, iniciace pohybů atd.
Syntéza a inaktivace acetylcholinu
Acetylcholin je ester cholinu a acetátu. Je tvořen jednokrokovou syntézou: přenos acetylu z AcCoA na cholin pomocí cholinacetyltransferázy. V synaptické štěrbině se nachází enzym acetylcholinesteráza (AChE), který štěpí acetylcholin na cholin (transport do presynaptického zakončení) a acetát. To vede k ukončení transmise.
Inhibitory AChE se uplatňují v terapii Alzheimerovy choroby a myashenia gravis; irreverzibilní inhibitory ze skupiny organofosfátů (využívané jako insekticidy) mohou způsobovat otravy.
Receptory pro acetylcholin
Acetylcholin působí skrze dva typy receptorů: muskarinové a nikotinové.
Muskarinové receptory
Metabotropní receptory spřažené s G-proteinem modulující činnost iontových kanálů. Celkem bylo popsáno 5 podtypů:
a) M1 receptory
„Neuronální” receptory vyskytující se převážně v CNS (kůra a hipokampální formace). Aktivace Gq-proteinu vede ke snížení efluxu K+ a excitaci. Jejich nedostatečná aktivita (či nižší hustota) podmiňuje demenci.
b) M2 receptory
Tzv. „kardiální” receptory, jsou exprimovány především v srdci, ale i v CNS. Spojené s Gi-proteinem – pokles cAMP vedoucí k otevření K+ kanálů, efluxu K+ a inhibici. Fungují jako autoreceptory (tlumí vyplavování ACh na synapsi) v kůře a hipokampu.
c) M3 receptory
Receptory „žláz a hladkých svalů”, spřaženy s Gq-proteinem, jejich aktivace vyvolává zvracení
d) M4 receptory
Působením i mechanismem účinku odpovídají M2-receptorům, vyskytují se převážně ve striátu.
e) M5 receptory
– spřaženy s Gq-proteinem (jako M1 a M3)
Nikotinové receptory
Ionotropní receptory – kanály propustné pro Na+, K+ a Ca2+. Existují dva základní podtypy:
muskulární (NM, lokalizován na nervosvalové ploténce) a
neuronální (NN, ve vegetativních gangliích).
NN-receptory fungují v CNS jako heteroreceptory jiných neurotransmiterových systémů (glutamátinergní, GABAergní, serotoninergní atd.) – podmiňují influx Ca2+ a zvyšují tak uvolnění neurotransmiteru.
Katecholaminy
Mezi katecholaminy řadíme noradrenalin (NA), dopamin (DA) a adrenalin. V CNS se uplatňují první dva.
Syntéza a inaktivace katecholaminů
Prekurzorem katecholaminů je aminokyselina L-tyrosin, jež je konvertována nejprv na L-dihydroxyfenylalanin (L-dopa). L-dopa je dále dekarboxylována na dopamin. Ten může být hydroxylován na noradrenalin, popř. dále methylován za vzniku adrenalinu. Ukončení neurotransmise je podmíněno zpětným transportem (reuptake) katecholaminů do neuronu a jejich zdejší inaktivací, kterou katalyzují dva enzymy – katechol-o-methyl transferáza (COMT) a monoaminooxidáza (MAO). Konečnými degradačními produkty katecholaminů jsou kyselina vanilmandlová a homovanilová.
L-dopa se využívá k terapii Parkinsonovy choroby – na rozdíl od hotových katecholaminů prostupuje L-dopa HEB, a může tak posílit tvorbu dopaminu, jež je u této choroby nedostatečná.
Noradrenergní systém
NA se uplatňuje jako modulátor ostatních neurotransmisních systémů, a to jak jako aktivátor, tak jako inhibitor (záleží na přítomných adrenergních receptorech). Noradrenergní neurony se nacházejí zejména v mozkovém kmeni, a to v locus coeruleus, tegmentu a retikulární formaci medully a pontu.
Axony neuronů z locus coeruleus projikují téměř do všech oddílů CNS, kde regulují funkce ostatních neurotransmisních systémů – modulují emoce, cyklus spánek-bdění či pozornost
Axony neuronů tegmenta a RF projikují do míchy, mozkového kmene, hypotalamu či limbického systému – ovlivňují řízení viscerálních funkcí, stresové reakce či excitabilitu míšních alfa-motoneuronů
Receptory pro noradrenalin
V CNS se vyskytují čtyři podtypy adrenergních receptorů: α1, α2, β1 a β2
1) α1-receptor a β1-receptor
Oba vykazují excitační účinky.
2) α2-receptor
Autoreceptor – presynaptická inhibice uvolňování NA.
3) β2-receptor
Vyskytuje se na gliových buňkách, jeho funkce není zcela objasněna.
Dopaminergní systém
Uplatňuje se v řízení motoriky, chování a viscerálních funkcí. Dopaminergní neurony jsou lokalizovány v mezencefalu a hypotalamu, můžeme popsat jejich čtyři základní projekce.
Dopaminergní projekce
1) Nigrostriatální projekce
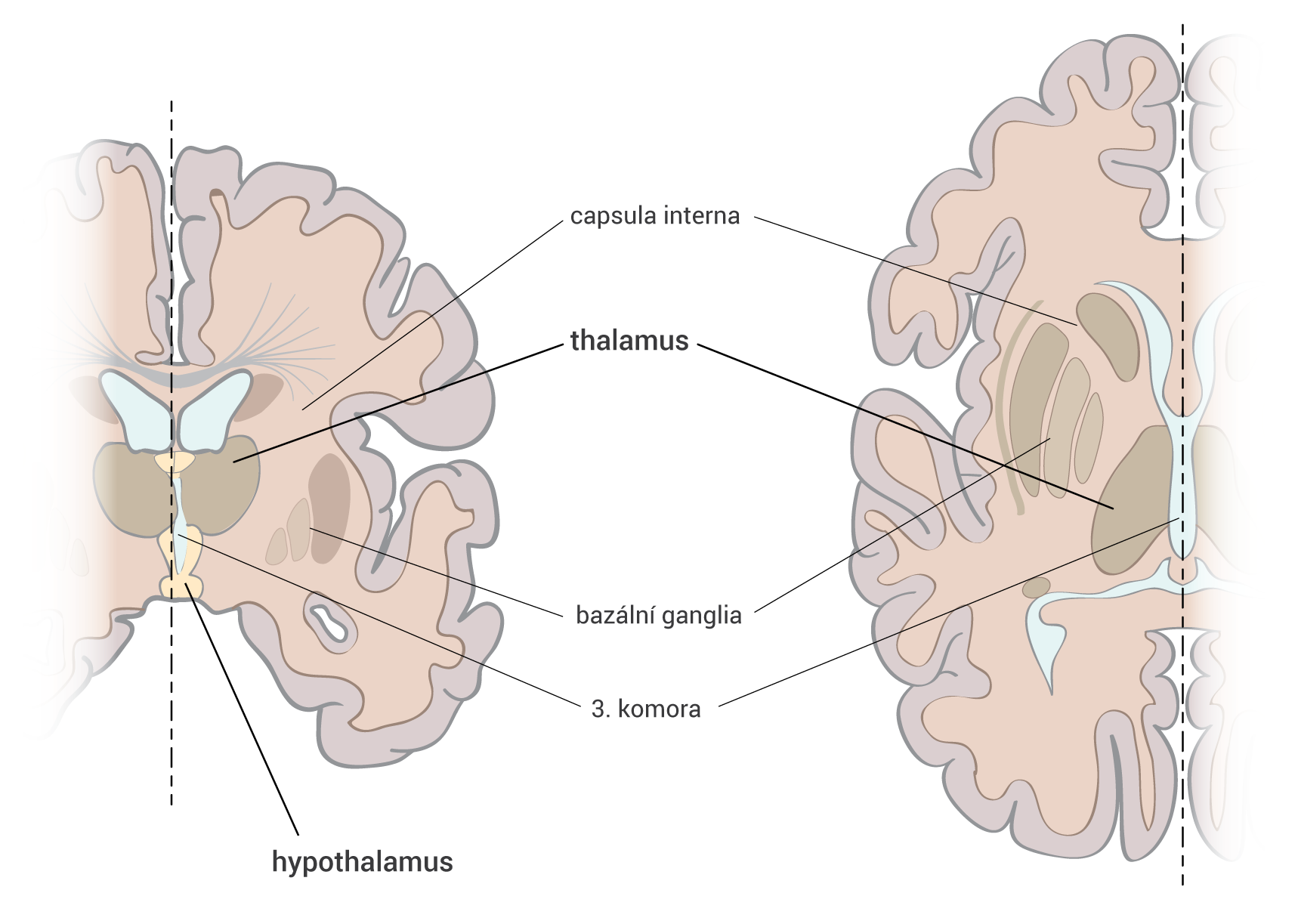
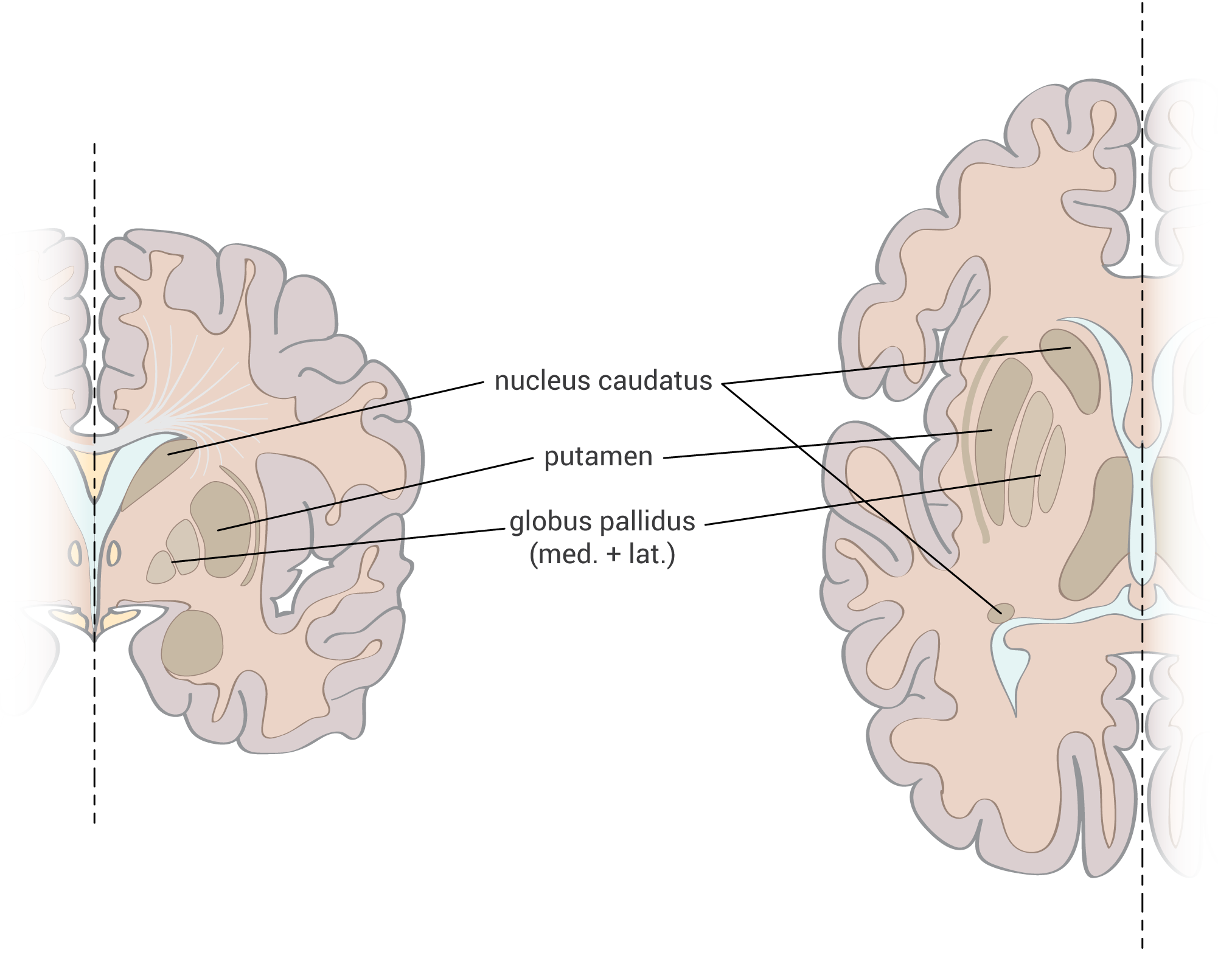
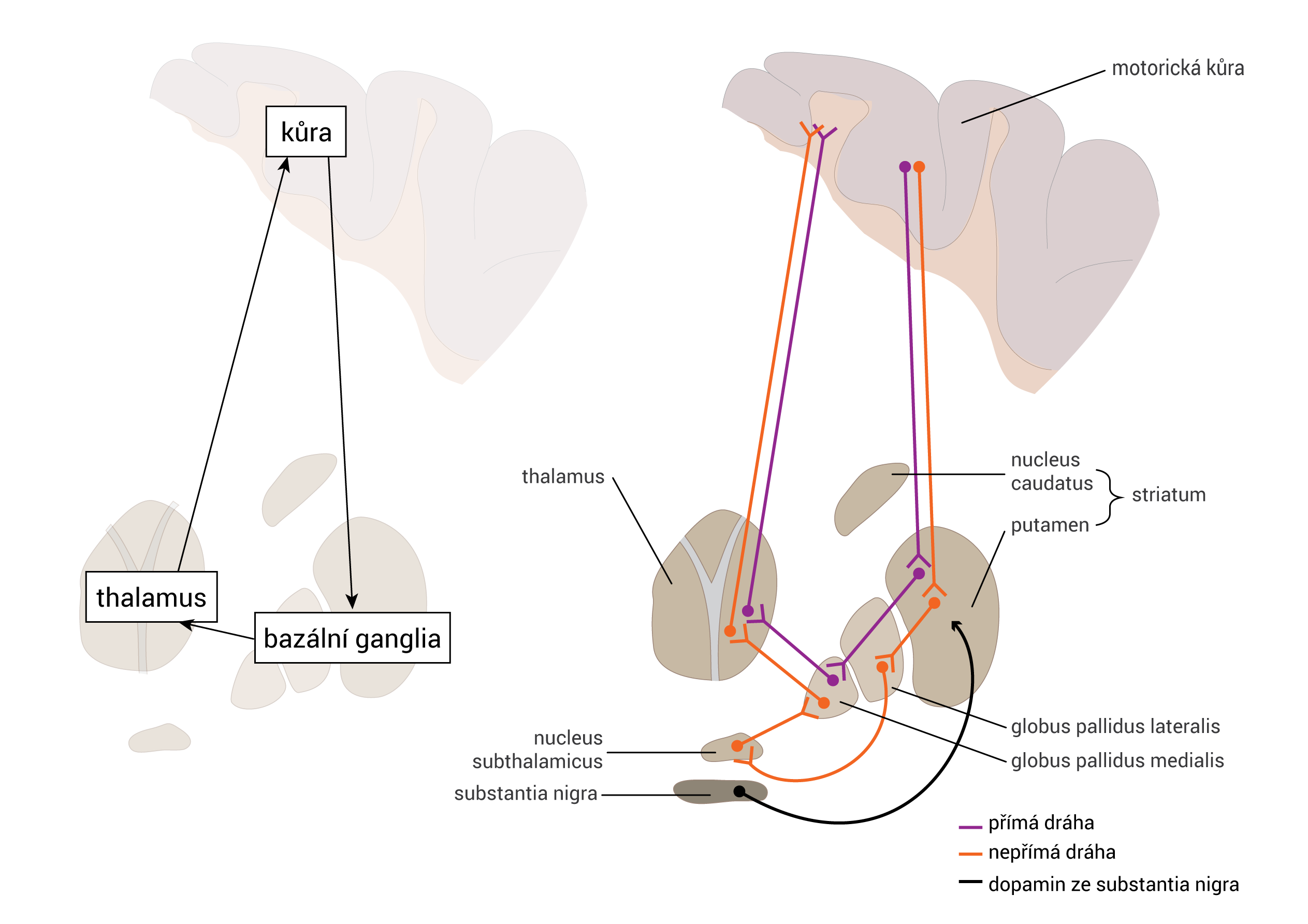
Axony začínají v substantia nigra a projikují do striata, globus pallidus a putamen – je zapojena do okruhu bazálních ganglií.
2) Mezolimbické projekce
Cílem neuronů z ventrálního tegmenta a mezencefalické RF jsou nc. accumbens, amygdala a hipokampus – hrají roli v motivaci, procesech odměny a trestu a vzniku závislostí
3) Mezokortikální projekce
Dalšími oblastmi, jež ovlivňují axony neuronů ventrálního tegmenta a mezencefalické RF jsou prefrontální oblasti kůry – modulují procesy frontálního kortexu, např. pozornost.
4) Tuberoinfundibulární projekce
Dopamin z hypothalamu inhibuje sekreci prolaktinu v adenohypofýze.
Receptory pro dopamin
Popsáno bylo pět typů receptorů, tzv. D1-5. Různá místa v CNS mají odlišné zastoupení jednotlivých typů receptorů: D2 v motorických oblastech kůry, D3 a D4 v limbickém systému.
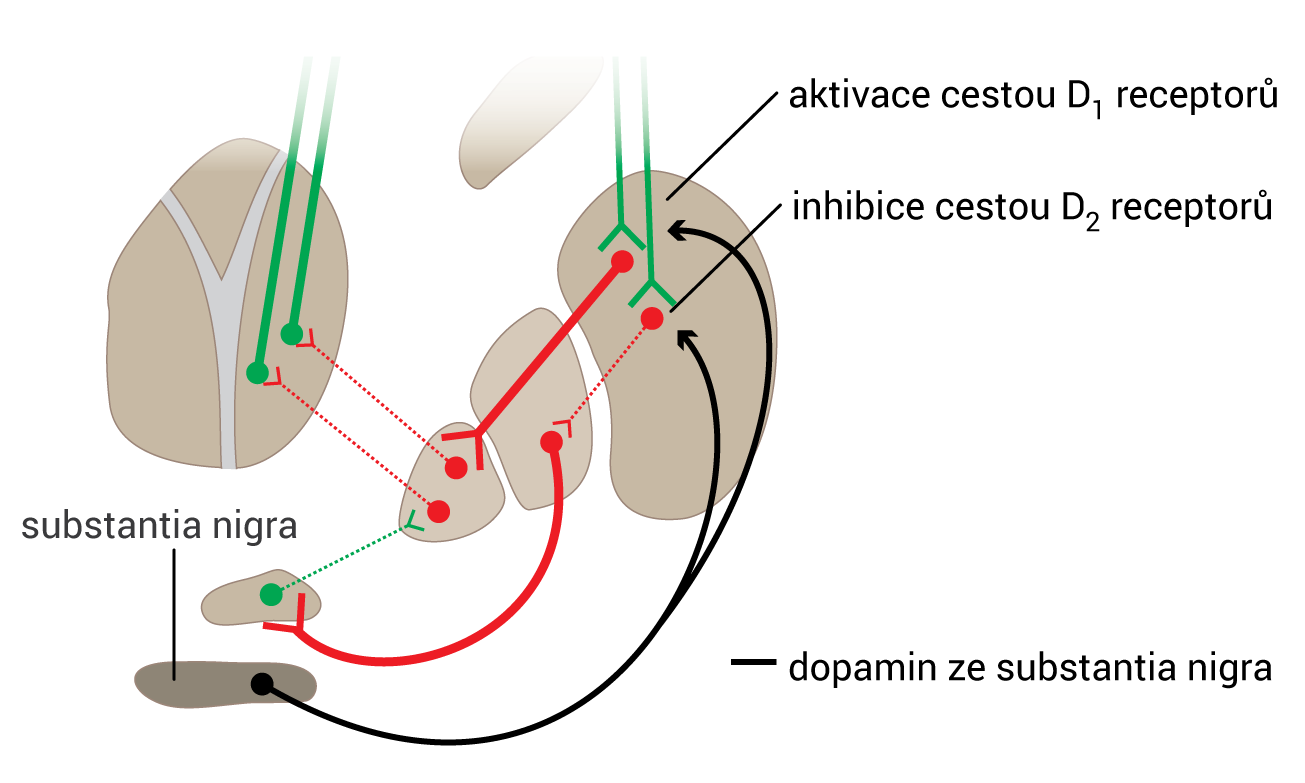
Všechny receptory jsou spřaženy s G-proteiny, jež modulují aktivitu adenylátcyklázy: D1 a D5 pomocí Gs-proteinu adenylátcyklázu stimulují; D2, D3 a D4 ji naopak pomocí Gi-proteinu inhibují.
Serotoninergní systém
Serotonin (5-hydroxytryptamin, 5-HT) moduluje aktivitu ostatních neurotransmisních systémů. Serotoninergní neurony se nacházejí v rafeálních jádrech RF – rostrální jádra projikují ascendentně, kaudální naopak descendentně.
1) Ascendentní serotoninergní systém (nc. raphe dorsalis, nc. raphe pontis centralis superior)
Projikuje do kůry, limbického systému, bazálních ganglií, hypothalamu i thalamu. Moduluje emoce, cyklus spánek-bdění či viscerální funkce.
2) Descendentní serotoninergní systém (nc. raphe pontis, nc. raphe magnus)
Vysílá eferenty ke kaudálním strukturám CNS: mozkovému kmeni, mozečku a míše. Modulace nocicepci (vnímání bolesti) i motoriku.
Syntéza a inaktivace serotoninu
Prekurzorem je aminokyselina L-tryptofan, z níž vzniká hydroxylací a následnou dekarboxylací. Ukončení neurotransmise probíhá reuptakem a inaktivací pomocí monoaminooxidázy, konečný degradační produkt serotoninu kyselina 5-hydroxyindoloctová.
Specifické inhibitory zpětného vychytávání serotoninu (SSRI) zvyšují aktivitu serotoninu na synapsi, čehož se využívá například v léčbě deprese.
Receptory pro serotonin
Existuje sedm podtypů serotoninergních receptorů, tzv. 5-HT1-7R: některé jsou excitační, jiné inhibiční – účinek serotoninu v různých oblastech je tedy podmíněn tamní expresí jednotlivých podtypů.
Glycinový systém
Glycin je inhibiční neurotransmiter lokalizovaný převážně v šedé hmotě míšní. Funguje prostřednictvím ionotropního receptor spřaženého s Cl- kanálem.
Histaminergní systém
Histamin je biogenní amin, vzniká dekarboxylací histidinu. Uplatňuje se v regulaci cyklu spánek-bdění a fungování hypotalamohypofyzárního systému.
Receptory pro histamin
Tři základní typy receptorů: H1-3
1) Receptory H1
Poklesem propustnosti membrány pro K+ vyvolávají depolarizaci a tedy excitaci. Vyskytují se v hypotalamu a mammilárních tělíscích. Léky na bázi antihistaminik vyvolávají únavu oslabením jejich činnosti.
2) Receptory H2
Receptory spřažené s Gs-proteinem a lokalizované na korových neuronech, gliích a endotelu.
3) Receptory H3
Autoreceptory tlumící vyplavování histaminu a heteroreceptory na NA, DA a ACh neuronech – tlumí uvolnění příslušného neurotransmiteru.
Peptidové neurotransmitery
Velmi široká skupina neurotransmiterů, řadíme mezi ně: opioidní substance (vazba na opioidní receptory, např. enkefaliny, endorfiny či dynorfin), substanci P, neuropeptid Y či somatostatin.
Syntéza peptidových NT
Tvorba probíhá v těle neuronu na ribosomech, hrubém endoplazmatickém retikulu a Golgiho aparátu; hotové peptidy jsou přeneseny axonálním transportem do presynaptického terminálu. Na ribosomech vznikají delší prekurzory (tzv. prepropeptidy), které se postupně zkracují (nejdříve na tzv. propeptidy) a přeměňují na funkční neurotransmiter. Pro některé neurotransmitery stojí na počátku velká mateřská molekula, jež je v několika krocích štěpena na více kratších aktivních neurotransmiterů. Mezi tyto mateřské molekuly patří proopiomelanokortin (POMC), jenž dá vznik:
- β-endorfinu,
- MSH (melanocyty stimulující hormon) a
- ACTH (adrenokortikotropní hormon)
Endogenní kanabonoidy
Patří mezi ně anandamid (arachidonoylethanolamid – AEA), derivát kyseliny arachidonové a etanolaminu.
Receptory pro kanabinoidy
Tzv. kanabinoidové receptory CB1 a CB2.
1) CB1 receptory
Vyskytují se převážně v CNS, jedná se o metabotropní receptory spřažené s Gi-proteinem, fungují jako heteroreceptory GABAergních neuronů – presynapticky inhibují uvolňování GABA do synaptické štěrbiny. Stimulují chuť k jídlu (orexigenní působení) a centra odměny.
2) CB2 receptory
Jsou lokalizovány v PNS a imunitním systému, uplatňují se ve vnímání bolesti.